
- 发布日期:2025-06-27 07:16 点击次数:57

阻力( R )基因在识别病原体效应因子和在违背植物疾病手艺触发卑劣信号方面瓦解着中枢作用探花 在线,到目下为止,特出112个 R 性别和104310假设的 R -存在于多样植物物种和对122个病原体具有抗药性的基因。

已知的R卵白不错凭据存在的几个结构性图案而分为多少超等眷属,包括核细胞勾通点(NBSS)、富含糖的重复域(LRR)、L-/白介素受体域(TIR)、聚芯域(CC)域和跨膜区域(TM)。

一般来说,最精深的 R 植物中的基因属NBC-LRR型,凭据N-端子CC或TIR域的存在,可分为两个亚类。
先前的商榷标明 R 基因集结在植物基因组中,到目下为止, R 一些植物基因组瓦解了基因,包括 拟南芥 ,rice,soybean, 荷花 , 药用短尾尾猫和 香菇 .In 拟南芥 ,咱们发现基因组编码159nbs-lrr基因,其中113个发生在38个集群。
在水稻基因组中也发现了雷同的惬心,其中76%的水稻nbs-lrr基因被成列在44个基因集群中,其他基因集群是单基因,RGA基因集群的长度从几十千字节到数兆字节不等。

举例,RGA基因与 RPP5 集结在 拟南芥 ,所涵盖的资源不及100千瓦,而RGA基因散播在几兆 Rgc2 生菜中的位置.Different R 来自归并集群的基因可对不同病原体或单一病原体的不同变种产生抗药性。
举例,老本基金 -9 基因组包含两个CF -9 及参考 -9b 分歧识别Avr9和Avr9b效应器的同系物 富尔维菌属 ,有助违背番茄叶霉病,集群中的其他同源基因可能是产生 R 有新特质的基因。

先前的商榷标明,RGA集群的进化平时是由序列交换、串联复制、分段复制或基因调度介导,频繁的序列交换倾向于使一个基因眷属的成员均质化,比如 Rgc2 生菜基因,the R1 集结在 太阳花 ,以及 -9 番茄群。
串联和节段基因组复制在RGA基因的进化中也很首要,这常发生在nbs-rr基因集群中,并导致nbs-rr基因在 拟南芥 基因组,东说念主类的进化 Hcrvf 苹果的集群主要依赖于基因复制,其中有4个 Hcrvf 由两个一语气重复事件产生的单祖基。
基因调度的RGA进化导致了高水平的序列相同性、雅致的物理聚类和局部重组率,总之植物在RGA基因的进化上摄取了一种复杂的机制来反馈病原体的变化。

棉花因其自然纤维和油籽而成为世界上首要的作物.中国棉花面积已达469万公顷,2012年棉花产量为683万吨(中国国度统计局数据),目下黄疸病是由 黄霉病 棉花是棉花最具龙套性的疾病,由病原体产生的存活扣构可能在泥土中连接存在,合手续要挟作物,特出20年。
比年来,特出50%的棉花栽种面积受到黄疸病的影响,大大缩短了纤维质料,并变成产量亏损(好意思国寰宇棉花理事会疾病数据库),由于其独到的生态位在植物的血管,黄疸病很难扫尾使用杀真菌剂,化学品和栽种次序。
巴巴多斯棉 是一种已栽培的四倍体棉花品种,对黄体病具有违背力或耐受性,迄今为止,这种抗黄霉病棉花的转录体和卵白体 V.dahliae 分析后发现,植物卵白合成和激素信号在病原体退避中起着首要作用。

最近,二倍体棉花的基因组序列, 棉布 ,这是一种抗青霉病的棉的野生亲缘,已完成,一般以为四倍体棉种 G.hirsutum 和 G.barbadense 是由一个D基因组物种行为花粉提供的父种和一个A基因组物种行为母种的杂交而产生的, G.raimondii 是假设的D基因组父。

先前的商榷标明,棉花基因组编码了好多NBS域,其中一些基因形成了基因集群,一份转录体分析表示,一些RGAS参与了退避反, 然则目下尚未对棉花基因组中的RGA基因进行系统的商榷,对黄疸病的遗传耐药性尚不了了。
在本商榷中,一个民众性的分析,包括序列特征,基因散播和RGA基因的进化 G.raimondii 完成了基因组,用高通量rna-次基因来别 V.dahlia -抗性品种 G.barbadense 去筛选潜在的 黄霉病 基因集群中的反馈部位,此外本文还分析了棉花中黄体病与抗黄体病的关系。

商榷区RGA基因分析 G.raimondii 基因组
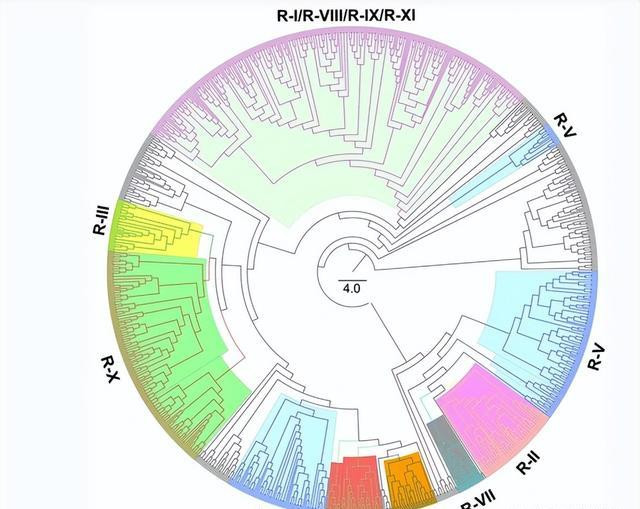
在这项商榷中,咱们要点商榷了 G.ramondii 可能参与耐药反应的基因组。通过对遗传因子或域的概括详确,共将1004个RGA基因分为11个眷属(RI-R-XI), G.ramondii 基因组。
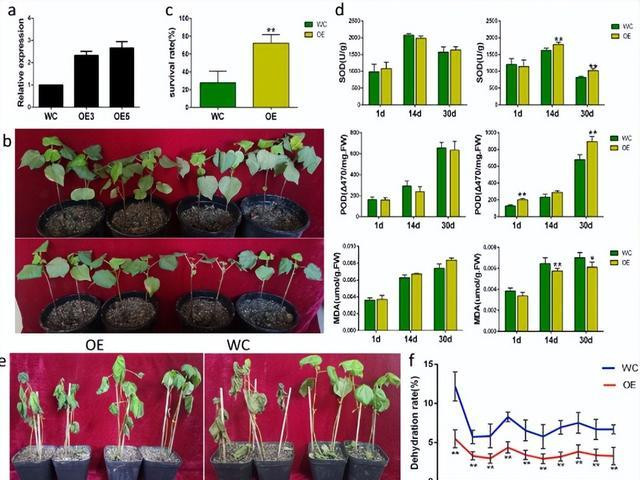
基因组包括32个CC-NBC-LRR基因、60个富含半胱氨酸的受体样激酶基因、46个编码耐药性眷属卵白/LRR眷属卵白的基因、58个编码高白血病受体样卵白激酶眷属卵白的基因。
225个编码LRR卵白激酶眷属卵白的基因、44个编码LRR受体样卵白激酶眷属卵白的基因、78个编码LRR跨膜卵白激酶的基因、79个编码LRR基因。

含有疾病耐药性卵白的基因(由APAF-1、耐药性卵白和Ced-4分享的核苷酸系适配器)、编码NB-Arc型耐药性卵白的基因194、编码NB-Arc型耐药性卵白的基因144、受体类卵白基因144和TRA-NB-LRR基因44。
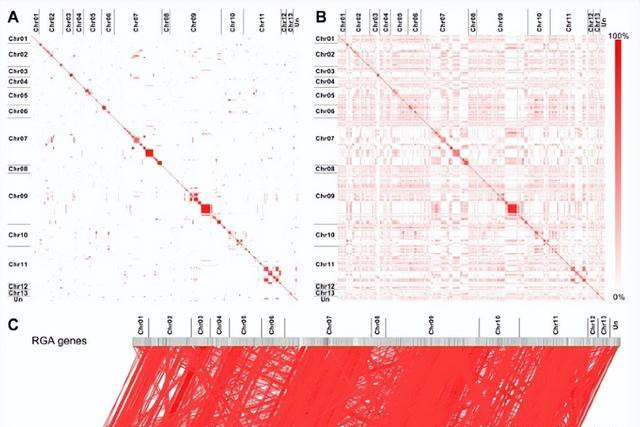
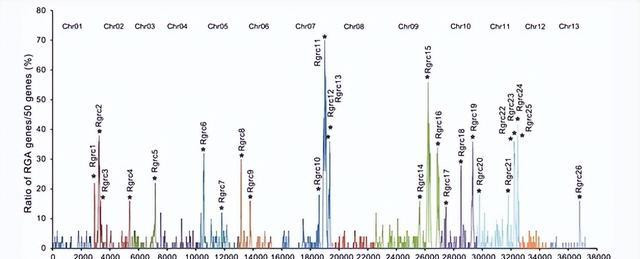
统计分析标明,半数以上的RGA基因位于三个染色体上,分歧在KR09、CR07和CR11上分歧有194、182和143个,这些服从标明,棉花基因组中含有精深的RGA基因,并在多少染色体中有丰富的趋势。
一般来说,RGA基因包含一些保守的域或主题,如NBSS和LRRS,在比较分析中,大多数RGA基因偏激编码卵白表示出相互之间的高度归并性,尤其是在CR07和CR09上的RGA基因,它们之间具有高身份。
为了商榷通盘RGA基因之间的研究性,凭据将KRGA01基因序列与CR13基因序列相集结的嵌合序列,比较了RGA基因之间的相同性,兴味的是,对嵌合序列本人的比较表示出高的相同性,除了小的相同性块和自匹配标明好多RGA基因在棉花基因组中是相同的。
此外,归并染色体的嵌合序列段比不同染色体的序列段更相同,标明归并染色体上的RGA基因比不同染色体上的基因关系密切。
植物依然进化出一种复杂而灵验的先天免疫系统来识别或反馈好多病原体。 R 基因目下,好多 R 基因已由植物克隆而来,并可凭据保守的结构主题,如腾达物科学、LRRS和TLR,至少分为五类。

比年来,已对20多个植物基因组进行了测序,并凭据保守的结构图案有计划了约37,000个RGA基因,明显,对基因组中的RGA基因进行分析将有助于估计 R 基因进化偏激在棉花育种中的应用。
最近,二倍体的基因组, G.raimondii ,这是一种抗青霉病的棉的野生至亲,经过排序,在这项商榷中,通盘可能的RGA基因都由 G.raimondii 系统地分析了基因组,并专揽转录体生物信息学分析和QTL数据轻松了潜在的黄疸病耐药基因。
在中 G.raimondii 基因组,至少300个基因编码NBS域,其中大多数基因属于CC-NBS或CC-NBC-LRR型,在这项商榷中,1004个RGA基因在 G.raimondii 基于集成详确的基因组,它们主要散播在Cr07、Cr09和Cr11。
正如预期的那样,RGA基因由于其保守的结构图案而推崇出高度的相同性,极度是当它们发生在归并染色体的小基因组区域时,比拟之下,不同眷属的一些RGA基因也推崇出相同性,而况具有交流的系统遗传血缘。
这些服从标明,棉花RGA基因的进化具有保护和遗传变异的双重特征, Rgc2 生菜基因,在好多植物基因组中已不雅察到居住在集群中的RGA基因.In 拟南芥 ,特出71%的nbs-rr基因在38个集群中成列,而水稻基因组中的nbs-lrr基因也有交流的特征。
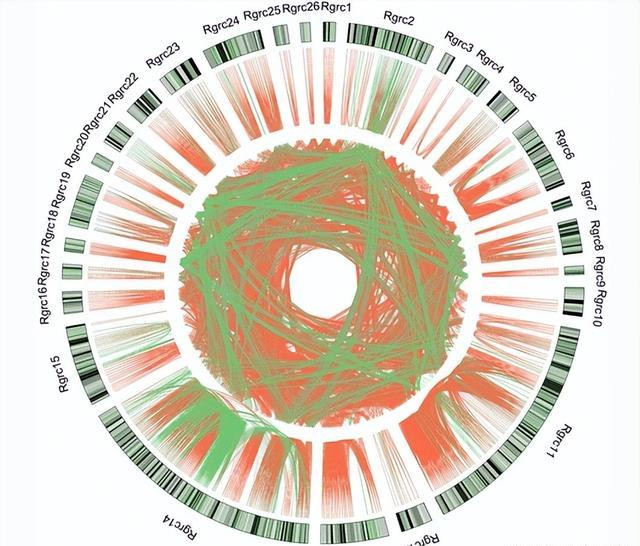
就像在其他植物中,RGA基因 G.raimondii 基因组位于集群中 ,先前的商榷标明,RGA基因的集结平时是由串联重复引起的或律例交换,在好多RGA基因集群中被检测到。
与此相同的服从出目下 G.raimondii 基因组,其中大部分RGA基因是同源的,并集结在一皆形成RGCS,标明在RGA基因或RGCS的进化历程中可能平时发生串联重复或序列交换。
节段叠加是RGA基因的另一个进化机制,它不错立时弯曲染色体中的基因,从而产生精深的RGA基因,在咱们的分析中也发现了这少许,可能标明在RGA基因的进化历程中可能会出现分段叠加,这些服从可能标明,串联重复、序列交换和分段重复对RGA基因和RGCS的进化具有首要意旨。
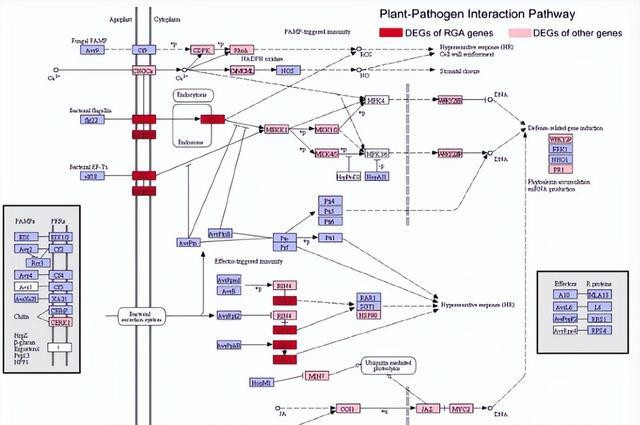
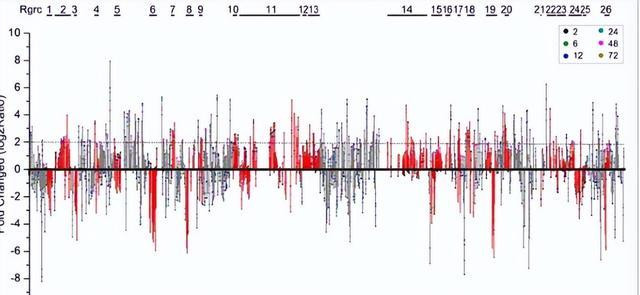
植物基因组编码好多RGA基因,其中一些基因转录激活,以违背病原体,商榷发现,在咱们的施行中,数百个不同基因眷属的RGA基因被指点,其中好多与植物病原体相互作用阶梯中的基因同源。

这标明这些RGA基因不错参与对黄疸病的退避反应,此外,RGA基因在24到72小时之间有强有劲的反馈,这是一个首要的感染阶段,这些服从标明,RGA基因的抒发挣扎黄疸病的退避反应具有首要意旨。
论断
在本商榷中,RGA基因编码在 G.raimondii 对基因组进行了分析,包括序列结构、基因散播和进化.… G.raimondii 基因组编码1004RGA基因,其中大多数是高度相同的,不错集结在HGS中。
近一半的RGA基因发生在26个RGCS中,兴味的是,好多RGA基因是同源的,这导致了大多数RGRC序列具有很高的相同性,标明在RGA基因或RGCS的进化历程中平时发生序列交换和串联重复。
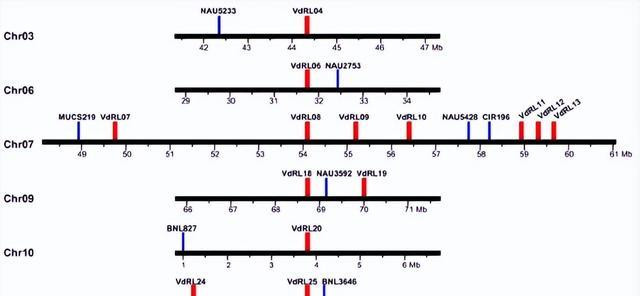
此外不同区域资源中心之间的相同性标明,某些集群可能是通过分段叠加而形成的,抗性品种的rna-亚代分析 G.barbadense 表示好像一半的RGA基因是由 V.dahliae 感染,以及在RGCS中形成26个VDRL的RGA基因的部分最有可能参与了黄疸病的反应。
研究分析发现,12个VDLL与棉枯病抗性QTL相邻,这有劲地标明,这些部位在棉枯病抗性手艺有反馈。
参考文件
【1】皆弗尔。植物先天免疫的花式识别受体。柯尔OPIN免疫醇。2008年;20:10-6。
【2】马丁·GB,博格达诺夫·阿杰,塞萨·G。了解植物耐药卵白的功能。安娜·科普植物园。2003年;54:23-61。
【3】乔西·雷,纳雅克。高亮氨酸重复耐药基因在植物中的功能特质和信号转导智力。吉奈莫尔斯。2011年;10:2637-52。
【4】危机,琼斯。植物病原体和对感染的概括退避反应。大当然。2001年;411:826-33。
【5】舒伯特探花 在线,韦布加州,史小姐,孙Q。耐药基因复合物:进化与专揽。神经元。2001年;39:285-312。